Vagal Stimulation and Arrhythmias
Chengzhe Liu1,2,3,4,, Hong Jiang1,2,3,4, Lilei Yu1,2,3,4,, Sunny S. Po5
1Department of Cardiology, Renmin Hospital of Wuhan University, Wuhan, China.2Cardiac Autonomic Nervous System Research Center of Wuhan Univer s ity, Wuhan, China.3Cardiovascular Research Institute, Wuhan University, Wuhan, China.4Hubei Key Laboratory of Cardiology, Wuhan, China.5Heart Rhythm Institute and Department of Medicine, University of Oklahoma Health Sciences
Center, Oklahoma City, O K USA.
I mbalance of the sympathetic and parasympathetic nervous systems is probably the most prevalent autonomic mechanism underlying many a rrhythmias . Recently, vagus nerve stimulation ( VNS has emerged as a novel therapeutic modality to treat arrhythmias through its anti adrenergic and anti inflammatory actions . C linical trials applying VNS to the cervical vagus nerve in heart failure pati en ts yielded conflicting results, possibly due to limited understanding of the optimal stimulation parameters for the targeted cardiovascular diseases. Transcutaneous VNS by stimulating the auricular branch of the vagus nerve, has attracted great attention d ue to its noninvasiveness. In this r eview, we summarize current knowledge about the complex relationship between VNS and cardiac arrhythmias and discuss recent advances in using VNS , particularly transcutaneous VNS , to treat arrhythmias.
Key Words : Vagus Nerve Stimulation, Autonomic Nervous System, Atrial Fibrillation, Ventricular Arrhythmia.
Correspondence to: Sunny S. Po, M.D., Ph.D
Section of Cardiovascular Diseases and
Heart Rhythm Institute
University of Oklahoma Health Sciences Center.
800 Stanton L Young Blvd, AAT 5400
, Oklahoma City, OK 73104, USA
Neuromodulation of the cardiac autonomic nervous system (ANS) is evolving as a novel approach to treat arrhythmias. Excitation of the parasympathetic nervous system exerts negative chronotropic, dromotropic and inotropic effects on the heart 1,2. Electrical stimulation of the vagus nerve (VNS) has emerged as a promising therapy for various conditions, including neural disorders and cardiac diseases 3-5. VNS was approved by the US Food and Drug Administration to treat refractory epilepsy and depression 6,7. Here, we focus on recent advances using VNS, particularly transcutaneous VNS, to treat arrhythmias.
1.1 Anatomy of the autonomic nervous system
Signal processing of the cardiac ANS occurs at several levels: i) central; ii) intrathoracic extracardiac; and iii) intrinsic cardiac level 8. Neural trafficking is influenced by the brain, spinal cord, extrinsic and intrinsic cardiac ganglia Figure 1. Autonomic neural signals from other organ systems (e.g. kidneys) can affect the cardiac ANS through complex interactions in the ANS 9-12.
Figure 1. Neurohumoral control and functional organization of cardiac autonomic innervation.
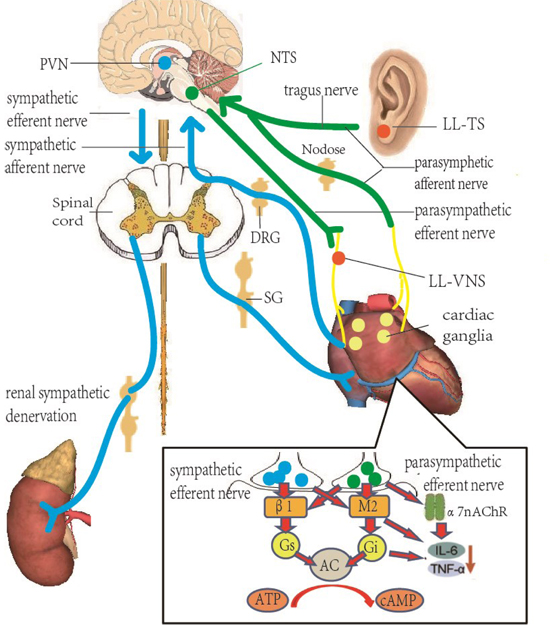
The autonomic nervous system related to arrhythmias consists of neurons and nerves in the brain, spinal cord, heart and kidneys and is similar to a closed-loop circuit that modulates the function of target organs. Activation of both the afferent and efferent vagal nerve fibers can increase the vagal tone in the cardiac closed-loop circuit and protect the heart. Blue lines represent sympathetic nerve fibers and green lines represent vagus nerve fibers. Inset: At the cellular level, sympathetic nervous system primarily releases norepinephrine, which stimulates the cardiac β-receptors. Parasympathetic nervous system primarily releases acetylcholine, stimulating cholinergic muscarinergic receptors on the myocytes and activating the α7nAChR pathway to reduce inflammation and fibrosis in the heart. SG, stellate ganglion; DRG, dorsal root ganglia; PVN, paraventricular nucleus; NTS, nucleus tractus solitaries; β1, β-adrenergic receptor; M2, muscarinic receptor; Gi, inhibitory G-protein; Gs, stimulatory G-protein; AC, adenylate cyclase; α7nAChR, α7 nicotinic acetyl-choline receptor.
1.2 Sympathetic Efferent Neurotransmission
The cardiac sympathetic preganglionic fibers originate in the central nervous system primarily in the brainstem and are modulated by higher centers such as the subthalamic and periaqueductal grey as well as the rostral ventrolateral medulla 9-12. Then, the sympathetic preganglionic fibers reach postganglionic neurons in the superior cervical, middle cervical, cervicothoracic (stellate) ganglia and mediastinal ganglia along the cervical and thoracic spinal cord (e.g. from C2 to T4 or T5) 9-12. These postganglionic neurons project axons via multiple cardiopulmonary nerves to the atrial and ventricular myocardium as well as limited populations of intrinsic cardiac adrenergic neurons. The major post-ganglionic neurotransmitter of the sympathetic nervous system is norepinephrine. The most important mechanism underlying sympathetic-mediated arrhythmogenesis is the activation of the β-adrenergic receptors and stimulatory Gs proteins, which leads to stimulation of adenylyl cyclase followed by protein kinase A–mediated phosphorylation of the L-type calcium channels (increasing calcium influx) and ryanodine receptors 13. Phosphorylation of the latter enhances the opening probability of the ryanodine receptors and increases calcium release from the sarcoplasmic reticulum (SR). Excessive calcium influx and SR calcium release are known to be arrhythmogenic because calcium homeostasis is crucial in maintaining normal cardiomyocyte functions such as excitability and mitochondrial stability. Elevated intracellular calcium concentration can activate the sodium-calcium exchanger (NCX) to extrude intracellular calcium to the extracellular space. However, extruding one calcium ion occurs at the expense of importing 3 sodium ions, which is electrogenic and can lead to early or delayed after-depolarization 14. Therefore, disturbed calcium homeostasis has been implicated as a leading mechanism underling high sympathetic outflow induced ventricular tachyarrhythmias (VAs) such as catecholaminergic polymorphic VT, long QT syndrome and heart failure.
1.3 Parasympathetic Efferent Neurotransmission
Preganglionic neurons of the parasympathetic nervous system are located in the nucleus ambiguous and dorsal motor nucleus of the medulla oblongata as well as scattered regions between these two structures 15,16. Their axons project to the postganglionic parasympathetic neurons in the numerous intrinsic cardiac ganglia via bilateral vagosympathetic trunks and multiple intrathoracic cardiopulmonary nerves 17. Postganglionic neurons, concentrated in epicardial fat pads, then provide direct innervation to the sinus node, atrioventricular node as well as both atria and ventricles 9,18,19. Acetylcholine is the major parasympathetic neurotransmitter of the heart; stimulation of the cholinergic muscarinic receptors (mainly the M2 receptors) inhibits adenylyl cyclase and reduces cyclic adenosine monophosphate via pertussis toxin-sensitive inhibitory G-proteins (Gi), which inhibits the L-type calcium current and hyperpolarization-activated current If as well as activates the Achgated potassium current (IKACh) 20. Important co-transmitters released with vagus nerve stimulation include nitric oxide and vasoactive intestinal peptide 21.
1.4 The intrinsic cardiac autonomic nervous system
Sympathetic and parasympathetic nerves and neurons as well as interconnecting nerves and neurons form a complex cardiac neural network. These neural elements converge at several ganglionated plexi (GP) embedded within epicardial fat pads 22,23. In the atria, the great majority of GP are concentrated at the pulmonary veinatrial junctions. In contrast, the ventricular GP are primarily located at the origins of major coronary arteries or aortic root 24. These GP act as integration centers that modulate the interactions between the extrinsic cardiac ANS and the heart 25 and contain both afferent and efferent sympathetic as well as parasympathetic nerves and neurons. For example, the bradycardic response elicited by cervical VNS was mediated by the anterior right GP adjacent to the sinus node; ablation of that GP greatly attenuated the bradycardic response 25.
1.5 Afferent Neurotransmission
Afferent nerve fibers from the mechanosensory and chemosensory receptors provide critical feedback from the cardiovascular system 26. Trafficking from these nerve fibers are processed in the intrinsic cardiac ganglia, intrathoracic ganglia, dorsal root ganglia of the spinal cord, nodose ganglia (the inferior ganglia of the vagosympathetic trunk) and brainstem 27. Afferent cardiac sympathetic neural trafficking is transmitted to the nucleus tractus solitaries (NTS) and the paraventricular nucleus (PVN) 28-31. In addition to projections from the PVN to the neurohypophysis, anatomic and electrophysiological studies revealed that axons from the PVN also project directly to the autonomic centers in the medulla and spinal cord, indicating that the PVN is a key integrative center for the sympathetic neural trafficking in the brain and is involved in cardiovascular regulation 32,33. Parasympathetic afferent fibers carry peripheral information to the NTS first; axons from the NTS project to the autonomic and cardiovascular centers in the brainstem as well to the hypothalamus and cerebrum. It is important to note that the afferent parasympathetic neural trafficking from peripheral organs back to the brain allows the brain to modulate the ANS and maintain autonomic homeostasis.
2. Vagus nerve stimulation to treat atrial fibrillation
2.1 Rationale for vagus nerve stimulation to treat atrial fibrillation
Simultaneous recordings of the canine left stellate ganglion (LSG) and left vagus nerve over several weeks revealed that coactivation of the sympathetic and parasympathetic nervous systems may precede paroxysmal AF Figure 234,35. That is, sympathetic and parasympathetic activity act synergistically to facilitate AF initiation38,39. In isolated atrial myocytes, parasympathetic stimulation shortened the atrial effective refractory period (ERP), whereas sympathetic stimulation increases calcium influx and SR calcium release which activates NCX, depolarizes the myocytes and elicit and early after-depolarization 36,37. Parasympathetic stimulation activates acetylcholine dependent potassium currents (IKACh), leading to shortening the atrial ERP and action potential duration (APD) 20,40,41 as well as a reduction in the atrial reentrant wavelength (the product of ERP and conduction velocity) to increase the probability that multiple reentrant circuits coexist in the atrial myocardium and facilitate AF maintenance 42.
Figure 2. Simultaneous recording of ECG, stellate ganglion nerve activity (SGNA), vagus nerve activity (VNA) and superior left GP nerve activity (SLGPNA) in ambulatory dogs.
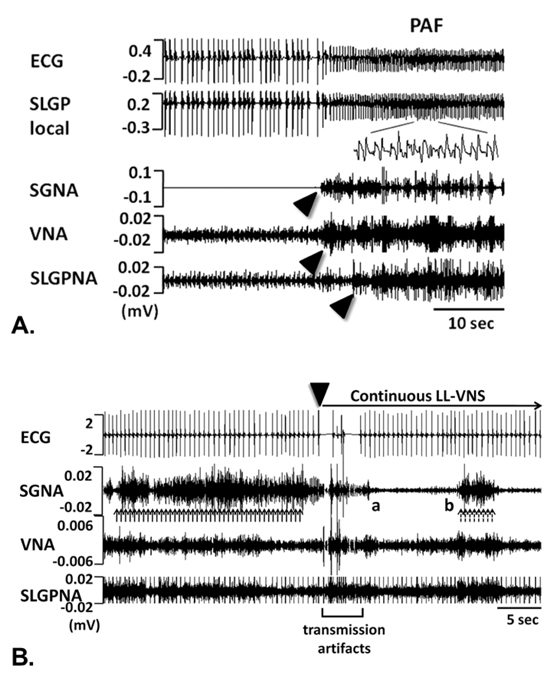
A. Paroxysmal AF was preceded by nearly simultaneous activation of the SGNA, VNA and LSLGPNA. B. LL-VNS immediately suppressed SGNA, demonstrating its anti-adrenergic effect. Reproduced with permission from reference
50.
Direct VNS produces atrial ERP heterogeneity due to the heterogeneous distribution of vagal innervation and varying density of the M2 receptors in the atria 43. In past decades, VNS, at the strength to slow the sinus rate or atrioventricular (AV) conduction, was used as an experimental tool to induce and maintain AF 44,45. In contrast, mild activation of vagal tone through the baroreflex has been shown to suppress firing of pulmonary veins 46. This paradox illustrates the complexity of the cardiac ANS and arrhythmogenicity. That is, VNS can either enhance or suppress AF, depending on the strength of stimulation 47.
2.2 Cervical low-level vagus nerve stimulation
The Oklahoma group first reported the antiarrhythmic effect of applying low-level VNS (LL-VNS) to canine cervical vagus nerve 48. LL-VNS, without slowing the sinus rate or AV conduction, increased the ERP in the atrium and pulmonary veins, suppressed AF inducibility, and shortened the duration of acetylcholine-induced AF 48,49. Since the atrial autonomic neural network is dominated by parasympathetic neural elements, inhibiting the GP by LLVNS leads to anti-cholinergic effects on GP and prolonged the ERP. Other mechanisms that LL-VNS suppresses AF have been proposed, including suppression of the LSG activity 50, release of the neurotransmitter vasostatin-1 51 and nitric oxide 52. Direct neural recordings of the canine atrial GPs showed that LL-VNS could inhibit the neural activity of GPs, thereby suppressing AF 49. Studies on ambulatory dogs demonstrated that paroxysmal AF was often initiated by simultaneous or sequential firing of the stellate ganglion, vagus nerve and GP Figure 2. LL-VNS inhibited the LSG activity and sympathetic nerve density in the LSG, thereby suppressing paroxysmal atrial tachyarrhythmias 53. These findings indicated that LL-VNS was both anticholinergic and antiadrenergic, which may account for its antiarrhythmic effects.
High sympathetic outflow enhances inflammation; inflammation leads to fibrosis through activation of pro-inflammatory cells (e.g. T-lymphocytes, monocytes/macrophages) and the cytokines they release. Inflammation, therefore, plays an important role in the pathogenesis of AF as well as neural, electrical and structural remodeling 54. Since the discovery of the α7 nicotinic acetylcholine receptor (α-7nAChR)-mediated cholinergic anti-inflammatory pathway, the anti-inflammatory effects of the parasympathetic nervous system on cardiovascular diseases have attracted substantial attention 55. Some studies suggested that activation of α7nAChR significantly reduces inflammation and fibrosis in the heart, in which the expression levels of high-mobility group box 1 (HMGB1), chemokine receptors and pro-inflammatory factors such as interleukin-6 and TNF-∝ were decreased56. In an ischemia/reperfusion model, VNS increased STAT3 phosphorylation and inhibited NF-kB activation. The cholinergic anti-inflammatory pathway was involved in these effects 57.
Pre-clinical evidence indicates that LL-VNS is anti-arrhythmic and anti-inflammatory. Because of the invasive nature of cervical LLVNS, it has only been tested acutely in post-operative AF in patients undergoing open heart surgery. The incidence of postoperative AF was reduced by 66% by LL-VNS (20 Hz) for 72 hours after cardiac surgery 58.
2.3 Transcutaneous low-level vagus nerve stimulation
A major drawback of cervical LL-VNS is its invasiveness, requiring surgical implantation of a neurostimulator and a cuff electrode around the cervical vagus nerve. Adverse effects include infection, Horner syndrome, discomfort and pain at implant site 59-62. These adverse effects led to the investigation of transcutaneous LL-VNS. Tragus, a small pointed eminence of the external ear, is innervated by the auricular branch of the vagus nerve. The tragus is easily accessible to transcutaneous LL-VNS. Prior research using horseradish peroxidase to trace the cranial projection of the auricular branch of the vagus nerve found that the vagal afferent nerve fibers of the auricular branch terminate mainly in the NTS 63. It is important to note that VNS through the tragus only activates the afferent vagal neural trafficking because there is no efferent vagus nerves in the tragus that innervates the heart.
Preclinical studies showed that low-level tragus stimulation (LLTS), at the strength not slowing the sinus rate or AV conduction, exerted similar electrophysiological effects to cervical LL-VNS in terms of lengthening the ERP, suppressing pulmonary vein firing and AF as well as inhibiting the neural activity of major atrial GPs Figure 364,65. Notably, the anti-arrhythmic effects of LL-TS were still profound at the stimulation strength 80% below the threshold that slowed the sinus rate or AV conduction, suggesting that this level of stimulation might be tolerable in ambulatory patients with arrhythmias.
Figure 3. Effects of transcutaneous low-level vagus nerve stimulation on effective refractory period of atria, pulmonary veins and on neural activity of ganglionated plexi.
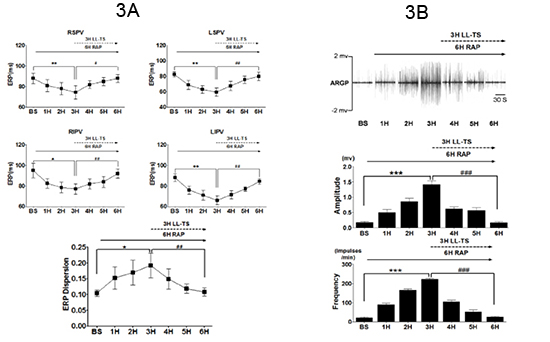
A. Parameters were measured during 6 hours of rapid atrial pacing (RAP) simulating paroxysmal AF. In the last 3 hours, LL-TS, 80% below threshold, was applied with RAP. At all sites, mean ERP decreased significantly after 3 hours of RAP (*:p<0.05; **:p<0.01; compared to baseline). After 3 hours of RAP+LL-TS, mean ERP at all sites showed a significant reversal toward baseline values (#:p<0.05 , ##:p<0.01; compared with the end of 3rd hour of RAP). Increased ERP dispersion by RAP was also reversed by LL-TS. B. Top. A typical example of neural recordings from the anterior right ganglionated plexi (ARGP) taken each hour (during sinus rhythm) when RAP was temporarily stopped. The middle and bottom panels showed the average amplitude and frequency of neural recordings. During the first 3 hours of RAP, there was a progressive increase in both the amplitude as well as the frequency of neural firing in the ARGP. With the addition of LL-TS, at 80% below threshold, the amplitude and frequency returned toward initial levels. RSPV, LSPV, RIPV and LIPV: right superior, left superior, right inferior and left inferior pulmonary vein, respectively. Reproduced from reference 65 with permission.
Electrical stimulation of the tragus was tested in 48 healthy participants showing that tragus VNS significantly decreased the low-frequency/high-frequency ratio (LF/HF) measurement of heart rate variability, indicating a tendency toward parasympathetic tone 66. In 2015, the Oklahoma group 67 reported a randomized clinical study applying transcutaneous LL-TS to patients with refractory paroxysmal AF referred for catheter ablation. Only one hour of transcutaneous LL-TS was enough to suppress ERP shortening and AF inducibility, shorten the AF duration, and decrease proinflammatory markers such as tumor necrosis factor-∝ (TNF-∝) and C-reactive protein. A recent sham-controlled randomized clinical trial published by same group indicated that in ambulatory patients with paroxysmal AF, daily transcutaneous LL-TS (one hour, 20 Hz, 1 mA below the perception threshold) reduced the AF burden by 83% at 6 months Figure 4. Plasma level of the TNF-∝ was reduced by 23% as well. These results suggest that transcutaneous LL-TS may serve as a novel, non-invasive therapy for patients in early stage of AF 68. However, as a major limitation of transcutaneous LL-TS, the response to transcutaneous LL-TS was variable among individual patients due to the lack of an acute biomarker of response to therapy that can predict the response to chronic transcutaneous LL-TS therapy Figure 4B. Although transcutaneous LL-TS has been shown to be able to affect heart rate variability and inflammatory markers within an hour 67, if these biomarkers predict long-term success remains unknown. Future large scale randomized clinical trials will be needed to optimize patient selection for transcutaneous LL-TS based on biomarkers as well as to determine if patients with more advanced stage of AF (e.g. persistent AF) still respond to transcutaneous LL-TS.
Figure 4. Effects of chronic transcutaneous low-level vagus nerve stimulation in atrial fibrillation burden.
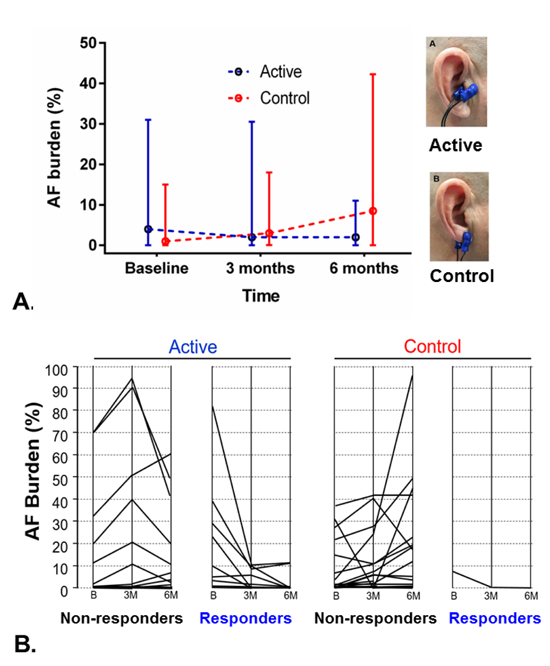
A. Comparison of AF burden between the 2 groups (median values and interquartile range). The p-value is based on a comparison of median AF burden levels at the 6-month time point after adjusting for baseline measures. In the control group, stimulation was delivered to the ear lobule where no vagal innervation exits. B. Patient-level data on AF burden change in the 2 groups. Patients whose AF burden decreased by >75% at follow up were categorized as responders. The proportion of responders was significantly larger in the active compared to the sham control group (47% vs. 5%, respectively, p=0.003). B = baseline; 3M = 3 months; 6M = 6 months. Reproduced from reference 68 with permission.
3. Vagus nerve stimulation to treat ventricular tachyarrhythmias
3.1 Rationale for VNS to treat ventricular tachyarrhythmias(VAs)
VAs are often triggered by high sympathetic tone or reduced vagal tone 69. Sympathetic activation can facilitate the initiation of VAs through the following mechanisms: 1) shortening of the ventricular ERP 70 and increasing the steepness of the slope of the action potential duration restitution curve to facilitate ventricular fibrillation initiation 71; 2) increasing dispersion of refractoriness 72; 3) enhancing of ventricular repolarization heterogeneity 73; and 4) triggering of early and delayed after-depolarization Figure 5) 14, 74, 75,. Furthermore, underlying cardiomyopathy can enhance the sympathetic activity and further promote the occurrence of VAs, forming a vicious cycle between the sympathetic activity and VAs. For instance, in a canine model of myocardial infarction, LSG synapses and nerve density were increased due to ischemia, which in turn caused more instability in the electrophysiological properties and increase the propensity for VAs 76.
Figure 5. Arrhythmogenesis related to high sympathetic outflow.
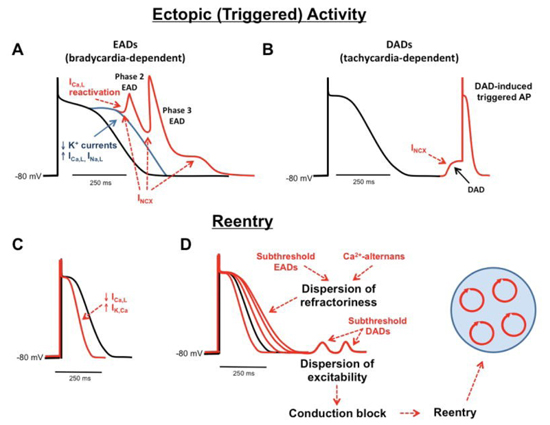
Both early afterdepolarization (EAD), A) and delayed afterdepolarization (DAD, B) can be elicited by the inward current generated by sodium-calcium exchanger (NCX). Reentry can be facilitated by shortened refractory period or action potential duration (C) as well as increased dispersion of the refractory period (D). Reproduced with permission from reference
14.
The beneficial effects of VNS on VAs are mediated directly by reducing the sympathetic activity and indirectly by inhibiting myocardial remodeling and inflammation 61-62,77. Activation of the IKACh current through the muscarinic receptors and augmentation of neuronal nitric oxide production also contributes to the beneficial effects of VNS 78-81. In addition, inflammatory pathways have an important role in fibrosis 82, scar formation and hypertrophy 83; inflammatory mediators such as interleukin-1 can be directly arrhythmogenic 84. In a rat model of ischemia/reperfusion, VNS reduced the infarct size, inflammatory cell infiltration and the levels of circulating inflammatory cytokines 85. Chronic VNS in a dog model of heart failure also normalized the levels of interleukin-6 and TNF-∝86and reduced plasma levels of angiotensin-II 87, a potent profibrotic mediator. Moreover, chronic VNS preserved the connexin 43 proteins and reduced the prevalence of spontaneous ventricular tachycardia after myocardial infarction 88.
At present, clinical management of ventricular tachycardia/ ventricular fibrillation is often restricted to pharmacological therapy and catheter ablation. Lately, invasive procedures such as thoracic epidural anesthesia (TEA), stellate ganglion blockade and cardiac sympathetic denervation (CSD), aiming at decreasing sympathetic outflow to the heart, have been shown to reduce the incidence of VTs in various conditions 89,91. The use of TEA is limited by antiplatelet or anticoagulation therapy due to concerns about bleeding. The effect of stellate ganglion blockade as well as left CSD or bilateral CSD often depends on the operator; collateral damage to sympathetic innervation to the head, neck, and eyes can cause significant adverse effects 91-94.
3.2 Cervical low-level vagus nerve stimulation
Increased sympathetic tone is typical in patients with myocardial infarction or heart failure and is an important contributing factor to VAs. Preclinical studies demonstrated that VNS can increase ventricular electrical stability and protect against VAs during acute ischemia and reperfusion in animal models 95-99. Vanoli et al 100 showed that VNS effectively prevents ventricular fibrillation in conscious animals with myocardial infarction. During the repeated exercise stress tests, VNS decreased the incidence of ventricular fibrillation from 92% to 10%. Furthermore, VNS may stabilize the infarct border zones and reduce the incidence of VAs 101. Chen et al 102 found that LL-VNS with a stimulation voltage below the 80% voltage threshold required to slow the heart rate significantly decreased the incidence of VAs and exerts protective effects on myocardial ischemia/reperfusion injury, presumably by preserving the acetylcholine levels and intact parasympathetic neuronal pathways. At present, several clinical trials of VNS for the treatment of advanced heart failure have yielded conflicting results, probably caused by the combination of heterogeneous study population and lack of the knowledge of the optimal stimulation parameters 47,61,62.
3.3 Transcutaneous low-level vagus nerve stimulation
Due to the invasiveness of cervical LL-VNS, transcutaneous LLVNS has been investigated as a novel noninvasive method to treat VTs. Yu et al 103 found that in a canine post-myocardial infarction model, chronic transcutaneous LL-VNS (2h/day) for 2 months reduced inducibility of VTs, LSG neuronal activity, left ventricular remodeling and ANS remodeling at the infarct border zone. Recently, this group provided the first clinical evidence that when transcutaneous LL-VNS, 50% below the threshold slowing the sinus rate or AV conduction, was delivered at the time of ST elevation myocardial infarction, transcutaneous LL-VNS reduced the infarct size, myocardial ischemia/reperfusion related ventricular premature contraction and ventricular tachycardia as well as pro-inflammatory markers such as interleukin-1β, interleukin-6 and TNF-∝ in patients presenting with ST-segment elevation myocardial infarction undergoing percutaneous coronary intervention 104. This first-inman trial suggests that transcutaneous LL-VNS may be applied to patients in early stage of myocardial infarction to reduce myocardial injury and VAs.
While preclinical cervical VNS showed promising results in suppressing VAs, long-term beneficial outcomes have not been verified in clinical trials 61,62. For its noninvasiveness, transcutaneous LL-VNS is an attractive alternative to cervical VNS to treat VAs related to high sympathetic outflow such as ventricular tachycardia in patients with structural heart diseases and premature ventricular contraction. Future preclinical and clinical studies should focus on identifying the optimal stimulation parameters (e.g. frequency, pulse width, duty cycles) as well as acute biomarkers that can predict longterm efficacy.
Transcutaneous LL-VNS may offer a non-invasive and an inexpensive alternative to treat a variety of cardiovascular or inflammatory diseases related to high sympathetic outflow. The optimal stimulus parameters of VNS for individual disease are yet to be determined. Future pre-clinical and clinical studies are needed to clarify mechanisms responsible for its therapeutic effects and optimize the stimulation parameters fortargeted disease.
- Levy MN. Cardiac sympathetic-parasympathetic interactions. Federation proceedings. 1984; 43(11):2598-2602.
- Goldberger JJ, Arora R, Buckley U, Shivkumar K. Autonomic Nervous System Dysfunction. Journal of the American College of Cardiology. 2019; 73(10):1189-1206.
- Nahas Z, Teneback C, Chae JH, Mu Q, Molnar C, Kozel FA, Walker J, Anderson B, Koola J, Kose S, Lomarev M, Bohning DE, George MS. Serial vagus nerve stimulation functional MRI in treatment-resistant depression. Neuropsychopharmacology: official publication of the American College of Neuropsychopharmacology. 2007; 32(8):1649-1660.
- Ma J, Qiao P, Li Q, Wang Y, Zhang L, Yan LJ, Cai Z. Vagus nerve stimulation as a promising adjunctive treatment for ischemic stroke. Neurochemistry international. 2019; 131(null):104539.
- Spindler P, Bohlmann K, Straub HB, Vajkoczy P, Schneider UC. Effects of vagus nerve stimulation on symptoms of depression in patients with difficult-to-treat epilepsy. Seizure. 2019; 69(null) :77-79.
- Morris GL, Gloss D, Buchhalter J, Mack KJ, Nickels K, Harden C. Evidence-based guideline update: Vagus nerve stimulation for the treatment of epilepsy: Report of the Guideline Development Subcommittee of the American Academy of Neurology. Neurology. 81(16):1453-1459.
- Howland RH. Vagus Nerve Stimulation. Current Behavioral Neuroscience Reports. 2014; 1(2):64-73.
- Armour JA. Potential clinical relevance of the 'little brain' on the mammalian heart. Exp. Physiol. 2008; 93(2):165-176.
- Coote JH. Myths and realities of the cardiac vagus. Journal of Physiology. 2013; 591(17):4073-85.
- Fukuda K, Kanazawa H, Aizawa Y, Ardell JL, Shivkumar K. Cardiac Innervation and Sudden Cardiac Death. 2009; 5(4):289-295.
- Ardell JL, Andresen MC, Armour JA, Billman GE, Zucker IH. Translational Neurocardiology: preclinical models and cardioneural integrative aspects. J Physiol. 2016; 594(14):3877–3909.
- Kawashima T. The autonomic nervous system of the human heart with special reference to its origin, course, and peripheral distribution. Anatomy and Embryology. 2005; 209(6):425-438.
- Voigt N, Heijman J, Wang Q, Chiang DY, Li N, Karck M, Wehrens XHT, Nattel S, Dobrev D. Cellular and Molecular Mechanisms of Atrial Arrhythmogenesis in Patients With Paroxysmal Atrial Fibrillation. Circulation. 2014; 129(2):145-156.
- Landstrom AP, Dobrev D, Wehrens XHT. Calcium signaling and cardiac arrhythmias. Circ Res. 2017;120:1969-1993.
- Standish A, Enquist L, Schwaber J. Innervation of the heart and its central medullary origin defined by viral tracing. Science. 1994; 263(5144):232-234.
- Standish A, Enquist LW, Escardo JA, Schwaber JS. Central neuronal circuit innervating the rat heart defined by transneuronal transport of pseudorabies virus. 1995; 15(3 Pt 1):1998.
- Hopkins DA, Bieger D, Devente J, Steinbusch WM. Vagal efferent projections: Viscerotopy, neurochemistry and effects of vagotomy. 1996; 107(08):79.
- Rysevaite K, Saburkina I, Pauziene N, Vaitkevicius R, Noujaim SF, Jalife J, Pauza DH. Immunohistochemical characterization of the intrinsic cardiac neural plexus in whole-mount mouse heart preparations. Heart rhythm. 2011; 8(5):731-738.
- Brack KE, Coote JH, Ng GA. Quantitative analysis of the parasympathetic innervation of the porcine heart. Heart rhythm. 2010; 7(8):e2-e3, e3.
- Schotten U, Verheule S, Kirchhof P, Goette A. Pathophysiological mechanisms of atrial fibrillation: a translational appraisal. Physiol Rev. 2011; 91(1):265-325.
- Habecker BA, Anderson ME, Birren SJ, Fukuda K, Herring N, Hoover DB, Kanazawa H, Paterson DJ, Ripplinger CM. Molecular and cellular neurocardiology: development, and cellular and molecular adaptations to heart disease. Journal of Physiology. 2016; 594(14):3853-75.
- Armour JA, Murphy DA, Yuan BX, Macdonald S, Hopkins DA. Gross and microscopic anatomy of the human intrinsic cardiac nervous system. The Anatomical record. 1997; 247(2):289-298.
- Pauza DH, Skripka V, Pauziene N. Morphology of the Intrinsic Cardiac Nervous System in the Dog: A Whole-Mount Study Employing Histochemical Staining with Acetylcholinesterase. Cells Tissues Organs. 2002; 172(4):297-320.
- Armour JA. Functional anatomy of intrathoracic neurons innervating the atria and ventricles. Heart Rhythm. 2010; 7(7):994-996.
- Hou Y, Scherlag BJ, Lin J, Zhang Y, Lu Z, Truong K, Patterson E, Lazzara R, Jackman WM, Po SS. Ganglionated Plexi Modulate Extrinsic Cardiac Autonomic Nerve Input: Effects on Sinus Rate, Atrioventricular Conduction, Refractoriness, and Inducibility of Atrial Fibrillation. Journal of the American College of Cardiology. 2007; 50(1):61-68.
- Shivkumar K, Ajijola OA, Anand I, Armour JA, Chen PS, Esler M, De Ferrari GM, Fishbein MC, Goldberger JJ, Harper RM, Joyner MJ, Khalsa SS, Kumar R, Lane R, Mahajan A, Po SS, Schwartz PJ, Somers VK, Valderrabano M, Vaseghi M, Zipes DP. Clinical neurocardiology defining the value of neuroscience-based cardiovascular therapeutics. The Journal of physiology. 2016; 594(14):3911-3954.
- Armour JA, Huang MA, Pelleg A, Sylvén C. Responsiveness of in situ canine nodose ganglion afferent neurones to epicardial mechanical or chemical stimuli. Cardiovascular Research. 1994; (8):8.
- Affleck VS, Coote JH, Pyner S. The projection and synaptic organisation of NTS afferent connections with presympathetic neurons, GABA and nNOS neurons in the paraventricular nucleus of the hypothalamus. Neuroscience. 2012; 219(1-2):48-61.
- Xu B, Zheng H, Patel KP. Relative contributions of the thalamus and the paraventricular nucleus of the hypothalamus to the cardiac sympathetic afferent reflex. American journal of physiology. Regulatory, integrative and comparative physiology. 2013; 305(1):R50-R59.
- Wang WZ, Gao L, Pan YX, Zucker IH, Wang W. Differential effects of cardiac sympathetic afferent stimulation on neurons in the nucleus tractus solitarius. Neuroscience letters. 2006; 409(2):146-150.
- Wang WZ, Gao L, Pan YX, Zucker IH, Wang W. AT1 receptors in the nucleus tractus solitarii mediate the interaction between the baroreflex and the cardiac sympathetic afferent reflex in anesthetized rats. American journal of physiology. Regulatory, integrative and comparative physiology. 2007; 292(3):R1137-R1145.
- Yamashita H, Inenaga K, Koizumi K. Possible projections from regions of paraventricular and supraoptic nuclei to the spinal cord: electrophysiological studies.Brain Research. 1984; 296(2):373-378.
- Li YF, Jackson KL, Stern JE, Rabeler B, Patel KP. Interaction between glutamate and GABA systems in the integration of sympathetic outflow by the paraventricular nucleus of the hypothalamus. Ajp Heart & Circulatory Physiology. 2007; 291(6):H2847-H2856.
- Tan AY, Zhou S, Ogawa M, Song J, Chu M, Li H, Fishbein MC, Lin SF, Chen LS, Chen PS. Neural Mechanisms of Paroxysmal Atrial Fibrillation and Paroxysmal Atrial Tachycardia in Ambulatory Canines. Circulation. 2008; 118(9):916-925.
- Choi EK, Shen MJ, Han S, Kim D, Hwang S, Sayfo S, Piccirillo G, Frick K, Fishbein MC, Hwang C. Intrinsic Cardiac Nerve Activity and Paroxysmal Atrial Tachyarrhythmia in Ambulatory Dogs. Circulation. 2010; 121(24):2615-2623.
- Patterson E, Po SS, Scherlag BJ, Lazzara R. Triggered firing in pulmonary veins initiated by in vitro autonomic nerve stimulation. Heart rhythm. 2005; 2(6):624-631.
- Patterson E, Lazzara R, Szabo B, Liu H, Tang D, Li YH, Scherlag BJ, Po SS. Sodium-calcium exchange initiated by the Ca2+ transient: an arrhythmia trigger within pulmonary veins. Journal of the American College of Cardiology. 2006; 47(6):1196-1206.
- Shen MJ, Zipes DP. Role of the Autonomic Nervous System in Modulating Cardiac Arrhythmias. Circulation Research. 2014; 114(6):1004-1021.
- Oliveira M, Da Silva MN, Timoteo AT, Feliciano J, Sousa L, Santos S, Silva-Carvalho L, Ferreira R. Inducibility of atrial fibrillation during electrophysiologic evaluation is associated with increased dispersion of atrial refractoriness. Int. J. Cardiol. 2009; 136(2):130-5.
- Iwasaki Y K, Nishida K, Kato T, Nattel S. Atrial Fibrillation Pathophysiology Implications for Management. Circulation. 2011; 124(20):2264-2274.
- Wakili R, Voigt N, Kääb S, Dobrev D, Nattel S, Kääb S. Recent advances in the molecular pathophysiology of atrial fibrillation. Journal of Clinical Investigation. 2011; 121(8):2955.
- Allessie MA. Atrial Electrophysiologic Remodeling: Another Vicious Circle? J Cardiovasc Electrophysiol. 1999; 9(12):1378-1393.
- Arora R, Ulphani JS, Villuendas R, Ng J, Harvey L, Thordson S, Inderyas F, Lu Y, Gordon D, Denes P. Greene R, Crawford S, Decker R, Morris A, Goldberger J, Kadish AH. Neural substrate for atrial fibrillation: implications for targeted parasympathetic blockade in the posterior left atrium. Am. J. Physiol. Heart Circ. Physiol. 2008; 294(1): H134-44.
- Goldberger AL, Pavelec RS. Vagally-mediated atrial fibrillation in dogs: conversion with bretylium tosylate. International Journal of Cardiology. 1986; 13(1):47-55.
- Wang Z, Page P, Nattel S. Mechanism of flecainide\"s antiarrhythmic action in experimental atrial fibrillation. Circulation Research. 1992; 71(2):271-287.
- Tai CT, Chiou CW, Wen ZC, Hsieh MH, Tsai CF, Lin WS, Chen CC, Lin YK, Yu WC, Ding YA. Effect of phenylephrine on focal atrial fibrillation originating in the pulmonary veins and superior vena cava. Journal of the American College of Cardiology. 2000; 36(3): 788-793.
- Ardell JL, Nier H, Hammer M, Southerland EM, Ardell CL, Beaumont E, Kenknight BH, Armour JA. Defining the neural fulcrum for chronic vagus nerve stimulation: implications for integrated cardiac control. J Physiol. 2017; 595(22):6887-6903.
- Li S, Scherlag BJ, Yu L, Sheng X, Zhang Y, Ali R, Dong Y, Ghias M, Po SS. Low-level vagosympathetic stimulation: a paradox and potential new modality for the treatment of focal atrial fibrillation. Circulation. Arrhythmia and electrophysiology. 2009; 2(6):645-651.
- Yu L, Scherlag BJ, Li S, Sheng X, Lu Z, Nakagawa H, Zhang Y, Jackman WM, Lazzara R, Jiang H, Po SS. Low-Level Vagosympathetic Nerve Stimulation Inhibits Atrial Fibrillation Inducibility: Direct Evidence by Neural Recordings from Intrinsic Cardiac Ganglia. Journal of Cardiovascular Electrophysiology. 2011; 22(4):455-463.
- Shen MJ, Chang HC, Park HW, George Akingba A, Chang PC, Zhang Z, Lin SF, Shen C, Chen LS, Chen Z,Fishbein MC, Chiamvimonvat N, Chen PS. Low-level vagus nerve stimulation upregulates small conductance calcium-activated potassium channels in the stellate ganglion. Heart Rhythm.2013; 10(6):910-915.
- Stavrakis S, Scherlag BJ, Fan Y, Liu Y, Liu Q, Mao J, Cai H, Lazzara R, Po SS. Antiarrhythmic effects of vasostatin-1 in a canine model of atrial fibrillation. J Cardiovasc Electrophysiol. 2012; 23(7):771-777.
- Stavrakis S, Scherlag BJ, Fan Y, Liu Y, Mao J, Varma V, Lazzara R, Po SS. Inhibition of atrial fibrillation by low-level vagus nerve stimulation: the role of the nitric oxide signaling pathway. Journal of Interventional Cardiac Electrophysiology. 2013; 36(3):199-208.
- Shen MJ, Shinohara T, Park HW, Frick K, Ice DS, Choi EK, Han S, Maruyama M, Sharma R, Shen C, Fishbein MC, Chen LS, Lopshire JC, Zipes DP, Lin SF, Chen PS. Continuous low-level vagus nerve stimulation reduces stellate ganglion nerve activity and paroxysmal atrial tachyarrhythmias in ambulatory canines. Circulation. 2011; 123(20):2204-2212.
- Stavrakis S, Nakagawa H, Po SS, Scherlag BJ, Lazzara R, Jackman WM. The role of the autonomic ganglia in atrial fibrillation. JACC. Clinical electrophysiology. 2015; 1(null):1-13.
- Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, Li JH, Wang H, Yang H, Ulloa L, Al-Abed Y, Czura CJ, Tracey KJ. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature. 2003; 421(6921):384-388.
- Leib C, Goser S, Luthje D, Ottl R, Tretter T, Lasitschka F, Zittrich S, Pfitzer G, Katus HA, Kaya Z. Role of the Cholinergic Antiinflammatory Pathway in Murine Autoimmune Myocarditis. Circulation Research. 2011; 109(2):130-140.
- Zhao M, He X, Bi XY, Yu XJ, Gil Wier W, Zang WJ. Vagal stimulation triggers peripheral vascular protection through the cholinergic anti-inflammatory pathway in a rat model of myocardial ischemia/reperfusion. Basic research in cardiology. 2013; 108(3):345.
- Stavrakis S, Humphrey MB, Scherlag B, Iftikhar O, Parwani P, Abbas M, Filiberti A, Fleming C, Hu Y, Garabelli P, McUnu A, Peyton M, Po SS. Low-Level Vagus Nerve Stimulation Suppresses Post-Operative Atrial Fibrillation and Inflammation: A Randomized Study. JACC. Clinical electrophysiology. 2017; 3(9):929-938.
- Spuck S, Tronnier V, Orosz I, Schönweiler R, Sepehrnia A, Nowak G, Sperner J. Operative and Technical Complications of Vagus Nerve Stimulator Implantation. Neurosurgery. 2010;67(null): 489-94.
- Elliott RE, Morsi A, Kalhorn SP, Marcus J, Sellin J, Kang M, Silverberg A, Rivera E, Geller E, Carlson C. Vagus nerve stimulation in 436 consecutive patients with treatment-resistant epilepsy: Long-term outcomes and predictors of response. Epilepsy Behav. 2011; 20(1): 57-63.
- Premchand RK, Sharma K, Mittal S, Monteiro R, Dixit S, Libbus I, DiCarlo LA, Ardell JL, Rector TS, Amurthur B, KenKnight BH, Anand IS. Autonomic regulation therapy via left or right cervical vagus nerve stimulation in patients with chronic heart failure: results of the ANTHEM-HF trial. Journal of cardiac failure. 2014; 20(11):808-816.
- Zannad F, De Ferrari GM, Tuinenburg AE, Wright D, Brugada J, Butter C, Klein H, Stolen C, Meyer S, Stein KM, Ramuzat A, Schubert B, Daum D, Neuzil P, Botman C, Castel MA, D'Onofrio A, Solomon SD, Wold N, Ruble SB, Chronic vagal stimulation for the treatment of low ejection fraction heart failure: results of the NEural Cardiac TherApy foR Heart Failure (NECTAR-HF) randomized controlled trial. European Heart Journal. 2015; 36(7):425-433.
- Nomura S, Mizuno N. Central distribution of primary afferent fibers in the Arnold's nerve (the auricular branch of the vagus nerve): a transganglionic HRP study in the cat. Brain Res. 1984; 292(2):199-205.
- Yu L, Li X, Huang B, Zhou X, Wang M, Zhou L, Meng G, Wang Y, Wang Z, Deng J, Jiang H. Atrial Fibrillation in Acute Obstructive Sleep Apnea: Autonomic Nervous Mechanism and Modulation. J Am Heart Assoc. 6(9):e6264.
- Yu L, Scherlag BJ, Li S, Fan Y, Dyer J, Male S, Varma V, Sha Y, Stavrakis S, Po SS. Low-level transcutaneous electrical stimulation of the auricular branch of the vagus nerve: A noninvasive approach to treat the initial phase of atrial fibrillation. Heart Rhythm.2013; 10(3):428-435.
- Clancy JA, Mary DA, Witte KK, Greenwood JP, Deuchars SA, Deuchars J. Non-invasive Vagus Nerve Stimulation in Healthy Humans Reduces Sympathetic Nerve Activity. Brain Stimulation. 2014; 7(6):871-877.
- Stavrakis S, Humphrey MB, Scherlag BJ, Hu Y, Jackman WM, Nakagawa H, Lockwood D, Lazzara R, Po SS. Low-Level Transcutaneous Electrical Vagus Nerve Stimulation Suppresses Atrial Fibrillation. J. Am. Coll. Cardiol. 2015; 65(9): 867-75.
- Stavrakis S, Stoner JA, Humphrey MB, Morris L, Filiberti A, Reynolds JC, Elkholey K, Javed I, Twidale N, Riha P, Varahan S, Scherlag BJ, Jackman WM, Dasari TW, Po SS. TREAT AF (Transcutaneous Electrical Vagus Nerve Stimulation to Suppress Atrial Fibrillation): A Randomized Clinical Trial.JACC Clin Electrophysiol. 2020; 6(3): 282-291.
- Antzelevitch C. The Role of Spatial Dispersion of Repolarization and Intramural Reentry in Inherited and Acquired Sudden Cardiac Death Syndromes: John Wiley & Sons, Ltd; 2007.
- Martins JB, Zipes DP. Effects of sympathetic and vagal nerves on recovery properties of the endocardium and epicardium of the canine left ventricle. Circulation Research.1980; 46(1):100-110.
- Taggart, P. Effect of Adrenergic Stimulation on Action Potential Duration Restitution in Humans. Circulation. 2003; 107(2):285-289.
- Opthof T, Coronel R, Vermeulen JT, Verberne HJ, van Capelle FJ, Janse MJ. Dispersion of refractoriness in normal and ischaemic canine ventricle: effects of sympathetic stimulation. Cardiovascular Research. 1993;27(11):1954-60.
- Vaseghi M, Yamakawa K, Sinha A, So EL, Zhou W, Ajijola OA, Lux RL, Laks M, Shivkumar K, Mahajan A. Modulation of regional dispersion of repolarization and T-peak to T-end interval by the right and left stellate ganglia. Am. J. Physiol. Heart Circ. Physiol. 2013; 305(7):H1020-H1030.
- Ben-David J, Zipes DP. Differential response to right and left ansae subclaviae stimulation of early afterdepolarizations and ventricular tachycardia induced by cesium in dogs. Circulation. 1988; 78(5):1241-1250.
- Yu L, Huang B, Zhou X, Wang S, Wang Z, Wang M, Li X, Zhou L, Meng G, Yuan S. Renal sympathetic stimulation and ablation affect ventricular arrhythmia by modulating autonomic activity in a cesium-induced long QT canine model. Heart Rhythm. 2017; S1139054666.
- Cao JM, Fishbein MC, Han JB, Lai WW, Lai AC, Wu TJ, Czer L, Wolf PL, Denton TA, Shintaku IP, Chen PS, Chen LS. Relationship Between Regional Cardiac Hyperinnervation and Ventricular Arrhythmia. Circulation. 2000; 101(16):1960-1969.
- Vanoli E, De Ferrari GM, Stramba-Badiale M, Hull SS, Foreman RD, Schwartz PJ. Vagal stimulation and prevention of sudden death in conscious dogs with a healed myocardial infarction. Circulation Research.1991; 68(5):1471-1481.
- Ng GA, Brack KE, Patel VH, Coote JH. Autonomic modulation of electrical restitution, alternans and ventricular fibrillation initiation in the isolated heart. Cardiovascular Research. 2007; 73(4):750-760.
- Litovsky SH, Antzelevitch C. Differences in the electrophysiological response of canine ventricular subendocardium and subepicardium to acetylcholine and isoproterenol. A direct effect of acetylcholine in ventricular myocardium. Circulation Research. 1990; 67(3):615-627.
- Martins JB, Zipes DP, Lund DD. Distribution of local repolarization changes produced by efferent vagal stimulation in the canine ventricles. Journal of the American College of Cardiology. 1984; 2(6):1191-1199.
- Koumi S, Sato R, Nagasawa K, Hayakawa H. Activation of inwardly rectifying potassium channels by muscarinic receptor-linked G protein in isolated human ventricular myocytes. J Membr Biol. 1997; 157(1):71-81.
- Nicoletti A, Michel JB. Cardiac fibrosis and inflammation: interaction with hemodynamic and hormonal factors. Cardiovasc. Res. 1999; 41(3): 532-43.
- Klein R M , Vester E G , Brehm M U , Dees H, Strauer BE. [Inflammation of the myocardium as an arrhythmia trigger]. Zeitschrift für Kardiologie. 2000; 89 Suppl 3(3):24-35.
- De Jesus NM, Wang L, Lai J, Rigor RR, Francis Stuart SD, Bers DM, Lindsey ML, Ripplinger CM. Antiarrhythmic effects of interleukin 1 inhibition after myocardial infarction. Heart Rhythm. 2017; 14(5):727-736.
- Calvillo L, Vanoli E, Andreoli E, Besana A, Omodeo E, Gnecchi M, Zerbi P, Vago G, Busca G, Schwartz PJ. Vagal Stimulation, Through its Nicotinic Action, Limits Infarct Size and the Inflammatory Response to Myocardial Ischemia and Reperfusion. J Cardiovasc Pharmacol. 2011; 58(5):500-507.
- Wang H, Yu M, Ochani M, Amella CA, Tanovic M, Susarla S, Li JH, Wang H, Yang H, Ulloa L, Al-Abed Y, Czura CJ, Tracey KJ. Nicotinic acetylcholine receptor α7 subunit is an essential regulator of inflammation. Nature. 2003; 421(6921):384-388.
- Zhang Y, Popovic ZB, Bibevski S, Fakhry I, Sica DA, Van WagonerDR, Mazgalev TN. Chronic Vagus Nerve Stimulation Improves Autonomic Control and Attenuates Systemic Inflammation and Heart Failure Progression in a Canine High-Rate Pacing Model. Circ Heart Fail. 2009; 2(6):692-699.
- Ando M. Efferent Vagal Nerve Stimulation Protects Heart Against Ischemia-Induced Arrhythmias by Preserving Connexin43 Protein. Circulation. 2005; 112(2):164-170.
- Meissner A, Eckardt L, Kirchhof P, Weber T, Rolf N, Breithardt G, Van Aken H, Haverkamp W. Effects of thoracic epidural anesthesia with and without autonomic nervous system blockade on cardiac monophasic action potentials and effective refractoriness in awake dogs. Anesthesiology. 2001; 95(1):132-138, 6A.
- Kamibayashi T, Hayashi Y, Mammoto T, Yamatodani A, Taenaka N, Yoshiya I. Thoracic Epidural Anesthesia Attenuates Halothane-induced Myocardial Sensitization to Dysrhythmogenic Effect of Epinephrine in Dogs. Anesthesiology. 1995; 82(1):129-134.
- Bourke T, Vaseghi M, Michowitz Y, Sankhla V, Shah M, Swapna N, Boyle NG, Mahajan A, Narasimhan C, Lokhandwala Y, Shivkumar K. Neuraxial Modulation for Refractory Ventricular Arrhythmias: Value of Thoracic Epidural Anesthesia and Surgical Left Cardiac Sympathetic Denervation. Circulation. 2010; 121(21):2255-2262.
- Schwartz PJ, Periti M, Malliani A. The long Q-T syndrome. Circulation Arrhythmia & Electrophysiology. 2012; 5(4):868-877.
- Odero A, Bozzani A, De Ferrari GM, Schwartz PJ. Left cardiac sympathetic denervation for the prevention of life-threatening arrhythmias: The surgical supraclavicular approach to cervicothoracic sympathectomy. Heart Rhythm. 2010; 7(8):1161-1165.
- Schwartz PJ, Priori SG, Cerrone M, Spazzolini C, Odero A, Napolitano C, Bloise R, De Ferrari GM, Klersy C, Moss AJ, Zareba W, Robinson JL, Hall WJ, Brink PA, Toivonen L, Epstein AE, Li C, Hu D. Left Cardiac Sympathetic Denervation in the Management of High-Risk Patients Affected by the Long-QT Syndrome. Circulation. 2004; 109(15):1826-1833.
- Myers RW, Pearlman AS, Hyman RM, Goldstein RA, Kent KM, Goldstein RE, Epstein SE. Beneficial Effects of Vagal Stimulation and Bradycardia During Experimental Acute Myocardial Ischemia. Circulation. 1974; 49(5):943-947.
- Corr PB, Gillis RA. Role of the Vagus Nerves in the Cardiovascular Changes Induced by Coronary Occlusion. Circulation. 1974; 49(1):86-97.
- Kent KM, Smith ER, Redwood DR, Epstein SE. Electrical Stability of Acutely Ischemic Myocardium: Influences of Heart Rate and Vagal Stimulation. Circulation. 1973; 47(2):291-298.
- Kolman BS, Verrier RL, Lown B. The effect of vagus nerve stimulation upon vulnerability of the canine ventricle. American Journal of Cardiology. 1976; 37(7):1041-1045.
- Zuanetti G, De Ferrari GM, Priori SG, Schwartz PJ. Protective effect of vagal stimulation on reperfusion arrhythmias in cats. Circulation research. 1987; 61(3):429-435.
- Vanoli E, De Ferrari GM, Stramba-Badiale M, Hull SS, Foreman RD, Schwartz PJ. Vagal stimulation and prevention of sudden death in conscious dogs with a healed myocardial infarction. Circulation research. 1991; 68(5):1471-1481.
- Vaseghi M, Salavatian S, Rajendran PS, Yagishita D, Woodward WR, Hamon D, Yamakawa K, Irie T, Habecker BA, Shivkumar K. Parasympathetic dysfunction and antiarrhythmic effect of vagal nerve stimulation following myocardial infarction. JCI insight. 2017; 2(16):
- Chen M, Zhou X, Yu L, Liu Q, Sheng X, Wang Z, Wang S, Jiang H, Zhou S. Low-Level Vagus Nerve Stimulation Attenuates Myocardial Ischemic Reperfusion Injury by Antioxidative Stress and Antiapoptosis Reactions in Canines. Journal of cardiovascular electrophysiology. 2016; 27(2):224-231.
- Yu L, Wang S, Zhou X, Wang Z, Huang B. Chronic Intermittent Low-Level Stimulation of Tragus Reduces Cardiac Autonomic Remodeling and Ventricular Arrhythmia Inducibility in a Post-Infarction Canine Model. JACC: Clinical Electrophysiology. 2016.
- Yu L, Huang B, Po SS. Low-Level Tragus Stimulation for the Treatment of Ischemia and Reperfusion Injury in Patients With ST-Segment Elevation Myocardial Infarction A Proof-of-Concept Study. Jacc Cardiovascular Interventions. 2017; 10(15):1511.






